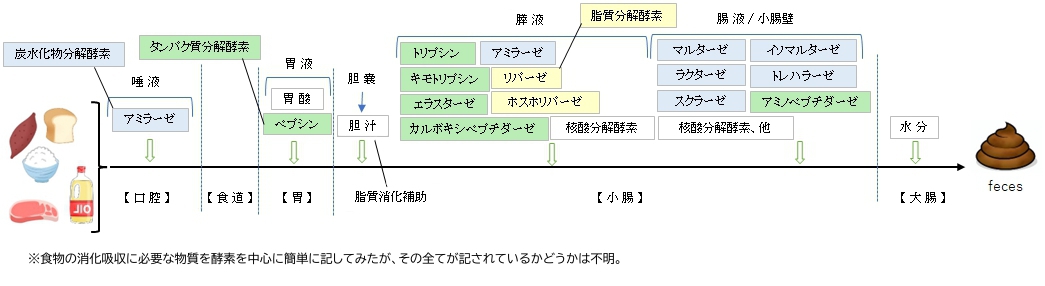
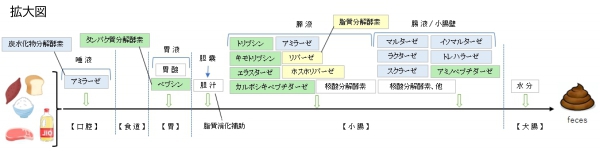
【概 要】

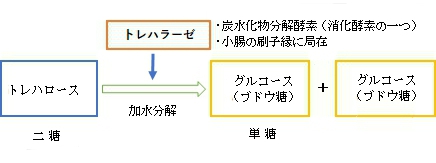
・「ほとんどの動物で見られる酵素であり、ヒトでは小腸の刷子縁に局在するグリコシダーゼである。」(ウィキペディア)
・「腸のトレハラーゼを欠くと、キノコなどのトレハロースを多く含むものを食べると、トレハロースが分解できないために吸収もできず、浸透圧性の下痢をする。」(ウィキペディア)
【タンパク質分解酵素一覧表】
|
唾液 |
アミラーゼ |
プチアリンとも。多糖である デンプンを主に二糖であるマルトースに分解する。 |
膵液 |
アミラーゼ |
膵アミラーゼ、アミロプシンとも。多糖である デンプンを主に二糖であるマルトース変える。 |
腸液 |
スクラーゼ |
二糖であるスクロースを ブドウ糖( 単糖)とフルクトース(果糖)に変える。 |
マルターゼ |
|
ラクターゼ |
二糖であるラクトース(乳頭)を 単糖である ブドウ糖とガラクトースに変える。 |
【参考となるサイト】
以下は「ウィキペディア」の解説文となる。
トレハラーゼ(Trehalase)とは、トレハロースを、グルコース2分子へと分解する反応を触媒する加水分解酵素である。ほとんどの動物で見られる酵素であり、ヒトでは小腸の刷子縁に局在するグリコシダーゼである。なお、トレハラーゼは1種類ではない。例えば、出芽酵母(Saccharomyces cerevisiae)は、最適pHの異なる2種類のトレハラーゼを持つ。中性トレハラーゼの最適pHは7.0、酸性トレハラーゼの最適pHは4.5である。出芽酵母の酸性トレハラーゼの活性の90%以上は細胞外であり、ペリプラズム内で細胞外のトレハロースをグルコースに分解していると報告されている。
トレハロースの分解
非還元性の二糖類であるトレハロース(α-D-グルコピラノシル-1,1-α-D-グルコピラノシド)は、重要な炭水化物の貯蔵形態の1つであり[注釈 1]、哺乳類を除く、地球上の多くの生物に存在する。1分子のトレハロースは、トレハラーゼによって2分子のグルコースに加水分解される[2][3][4][5]。トレハロースの加水分解は、1893年にエミール・ブルクロによってクロコウジカビ(Aspergillus niger)で初めて観察された。フィッシャーは1895年に出芽酵母でのこの反応を報告している。それ以来、トレハロースの加水分解酵素であるトレハラーゼは、植物や動物を含む多くの生物で報告されてきた。腸におけるトレハラーゼの機能は、経口摂取されたトレハロースを分解することである。腸のトレハラーゼを欠くと、キノコなどのトレハロースを多く含むものを食べると、トレハロースが分解できないために吸収もできず、浸透圧性の下痢をする。なお、トレハラーゼは腸以外でも発現していることがあり、トレハラーゼによるトレハロースの加水分解は、菌類の胞子の発芽、昆虫の飛行、休眠細胞の成長の再開など、多くの生物の生理的プロセスにとって重要である。
細菌のトレハラーゼ
シュードモナス属、バシラス属、リゾビウム属やその他放線菌目のいくつかは、トレハロースの形で糖を貯蔵しており、そして、恐らくその抵抗性に貢献していると報告されている。細菌から単離されるトレハラーゼの大部分の最適pHは、6.5から7.5である。Mycobacterium smegmatisのトレハラーゼは、膜結合タンパク質である。大腸菌(Escherichia coli K12)のペリプラズム内のトレハラーゼは、高い浸透圧の条件で生育することにより誘導される。トレハロースからグルコースへの分解はペリプラズム内で行われ、その後、グルコースは細胞内に取り込まれる。また、大腸菌には、細胞質に別のトレハラーゼが発現していることも報告されている。ただ、この細胞質内のトレハラーゼをコードする遺伝子は、ペリプラズムのトレハラーゼの遺伝子と高い相同性を示す。
植物のトレハラーゼ
トレハラーゼは植物に広く存在する。しかし、その基質であるトレハロースは植物に広く存在するとは言えない。植物界でトレハロースは、テマリカタヒバ(Selaginella lepidophylla)やヒメハナワラビ(Botrychium lunaria)などのシダ植物のいくつかの種に見られる。さらに、維管束植物ではトレハロースを持つ種は珍しく、セリ科のいくつかの種の熟した果実や、乾燥耐性被子植物であるミロタムヌス(Myrothamnus flabellifolius)の葉で報告されているのみである。
植物は基本的に光合成ができるため、グルコースやトレハロースを外部から摂取する必要性は無い。体内にもトレハロースを持たない植物、すなわち、トレハラーゼの基質が存在しない植物ですらトレハラーゼを持っている理由は謎であり、そのような植物中におけるトレハラーゼの活性に明確な役割は見い出せていない。しかし、トレハロースは植物の代謝を調節する機能を持つ前駆体のトレハロース-6-リン酸の合成を阻害する効果を示すため[6]、植物は何らかの理由で体内にトレハロースが入ってしまった場合には、そのトレハロースを除去する必要がある[7]。これらのことから、植物に発現しているトレハラーゼは、植物に関っている微生物が生産するトレハロースを分解する役割を持つと考えられている。
酵母のトレハラーゼ
出芽酵母では、少なくとも2種類のトレハラーゼが報告されている。1つは、cAMP依存性リン酸化により制御されていると報告されている。この酵素の活性は細胞質内で見られ、最適pHは約7.0であることから、中性トレハラーゼ(NT)と呼ばれる。もう1つのトレハラーゼ活性は液胞内で見られる。こちら酵素の最適pHは約4.5であることから、酸性トレハラーゼ(AT)と呼ばれる。これらの2つの酵素は、それぞれNTH1、NTH2という別の遺伝子にコードされている。
中性トレハラーゼ(NT)
中性トレハラーゼ(NT)は出芽酵母から単離され、同定された。非変性ゲル中では、このタンパク質の分子量は160 kDaを示すが、SDS-ポリアクリルアミドゲル電気泳動では80 kDaを示す。トレハロースに特異的な加水分解酵素で、Kmは5.7 mMと報告されている。S. cerevisiaeではNTH1遺伝子がコードしている。この遺伝子のオープンリーディングフレームは2079 bpで、693アミノ酸残基からなる79569 Daのタンパク質をコードしている。
NTの活性は、タンパク質のリン酸化-脱リン酸化により調節されている。cAMP依存性プロテインキナーゼによるリン酸化により、NTが活性化され、リン酸化された酵素をアルカリホスファターゼで脱リン酸化することにより酵素活性はほぼ完全に不活化される。アデノシン三リン酸及びプロテインキナーゼとともに置いて再リン酸化することにより、活性が復活することが観察されている。粗抽出物中のNTの活性はポリカチオンにより促進されるのに対して、リン酸化NTの活性はこれらにより阻害される。粗抽出物の活性化は、NT活性を阻害するポリリン酸の除去によることが分かっている。
酸性トレハラーゼ(AT)
酸性トレハラーゼ(AT)の分子量は、ゲルろ過クロマトグラフィーで218 kDaと測定されている。ATは糖タンパク質であり、炭水化物含量は86%である。ATの成熟化は、糖の付加のない41 kDaのタンパク質を出発として、まずは小胞体内で76 kDaの糖タンパク質となり、ゴルジ装置内でさらに糖化されて180 kDaになり、最終的に液胞内でさらに糖化されて約220 kDaになるという段階的な反応で行われる。純粋なATをエンドグリコシダーゼHで処理することで41 kDaの糖付加のないタンパク質が得られる。ATのトレハロースに対するKmは、pH4.5で約4.7 mMである。出芽酵母でATをコードする遺伝子は、ATH1である。
出芽酵母が細胞外のトレハロースを炭素源として利用するためには、Ath1p(AT)が必要である。ATH1を持たない突然変異体は、トレハロースを炭素源とする培地では生育できない。
ATは、合成地点からペリプラズムまで移動し、そこで外生のトレハロースに結合して内在化し、液胞内でそれを加水分解すると考えられている。出芽酵母のAT活性の90%以上が細胞外であり、ペリプラズムにおいてトレハロースをグルコースに分解していることも示されている。かつては、遺伝子YGP1の産物であり、高度に糖化されたタンパク質gp37がAT活性と関連していると報告されていた。またサッカラーゼの活性もAT活性と関連していると報告された。ATとこれら2つのタンパク質の物理的な関連は、Ath1pの既知の分泌シグナルの非存在下で、ATがサッカラーゼやgp37の分泌経路により分泌されると考えられる。