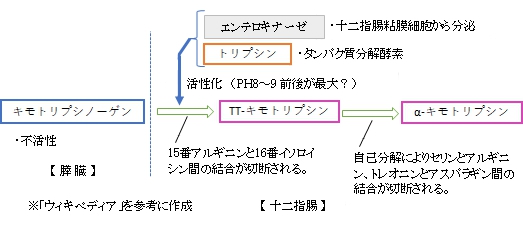
キモトリプシン( 英:chymotrypsin ) ・「エンドペプチダーゼ、セリンプロテアーゼの一種である。」(ウィキペディア) ・「芳香族アミノ酸のカルボキシル基側のペプチド結合を加水分解する。 」(ウィキペディア) ・「最適pHは8〜9程度の弱塩基性である。」(ウィキペディア) ・「生体内では、キモトリプシンはセリンプロテアーゼとして多くの臓器の消化系で働く。」(ウィキペディア) ・「」(ウィキペディア) 以下は「ウィキペディ」の解説文となる。 キモトリプシン(カイモトリプシン、chymotrypsin, EC.3.4.21.1・EC.3.4.21.2)はエンドペプチダーゼ、セリンプロテアーゼの一種である。膵液に含まれる消化酵素の一種で、芳香族アミノ酸のカルボキシル基側のペプチド結合を加水分解する。 膵臓からキモトリプシノーゲンとして分泌され、エンテロキナーゼ、トリプシンにより15番アルギニンと16番イソロイシン間の結合が切断されることにより、活性状態のπ-キモトリプシンとなる。その後、自己分解によりセリンとアルギニン、トレオニンとアスパラギン間の結合が切断され、α-キモトリプシンとなる。 遺伝子は第16染色体のq23-q24.1のCTRBである。 キモトリプシンが芳香族アミノ酸に対して基質特異性を発揮するのは活性中心の近辺に疎水性基でできた空洞があり、芳香族の側鎖がここに入ると安定化するためである。 ヒトではキモトリプシンの最適pHは8〜9程度の弱塩基性である。 酵素の反応メカニズムと反応速度 生体内では、キモトリプシンはセリンプロテアーゼとして多くの臓器の消化系で働く。この酵素は、触媒がない場合は熱力学的な安定性のためとてもゆっくりとしか起こらないペプチド結合の加水分解を促進する。この反応の基質(英語版)には、トリプトファン、チロシン、ロイシン、フェニルアラニンなどがあり、これらのアミノ酸のC末端を加水分解する。多くのプロテアーゼ同様、キモトリプシンも試験管内でアミド結合を加水分解するため、N−アセチル-L-フェニルアラニン-p-ニトロフェニルアミドのような反応物の類縁体の酵素分析(英語版)に用いられる。 キモトリプシンは反応性の低いカルボニル基を強力な求核剤である酵素の活性中心にある195番目のセリンの残基で攻撃し、一時的に基質と共有結合を作る。この時酵素基質複合体ができる。57番目のヒスチジン、102番目のアスパラギン酸のとともに195番目のセリンの残基は触媒三残基と呼ばれる。 これらの発見は抑制分析と、前述の基質の加水分解の反応速度の研究によるところが大きい。後者は、酵素基質複合体の4-ニトロフェノールが黄色い色をしており、410nmの吸光度を調べることで濃度が分かることが発見されたため可能になった。 キモトリプシンの基質との反応は2つの段階からなることがわかっている。初めは爆発的な段階で、次は安定な段階というミカエリス・メンテン式に従う反応である。またこれはピンポン機構とも呼ばれる。この反応は基質のアシル化が進み、アシル基 - 酵素複合体という反応中間体が形成される。その後、脱アシル化が進み、酵素は元に戻る。これは触媒三残基とともにある時に反応が起こる[1]。アスパラギン酸とヒスチジンのN-δ位の水素原子の間の水素結合によってε位の窒素原子のpKaが上がり、セリンが脱プロトン化できるようになる。このためセリンの側鎖が求核剤として働き、タンパク質の主鎖にある電子不足であるカルボニル基の炭素原子に結合することができるようになる。イオン化されたカルボニル基の酸素は主鎖の2つの水素原子による水素結合により安定化される。この反応はオキシアニオンホール(英語版)内で起こる。これにより四面体付加が起こり、ペプチド結合が開裂する。セリンに結合したアシル - 酵素複合体は、開裂して新たにできたタンパク質のN末端と結合し、セリンから離れる。反応の第2段階では、水分子が塩基性のヒスチジンに活性化されて求核剤として働くようになる。水分子中の酸素原子がセリンに結合したアシル基のうちのカルボニル基の炭素を攻撃すると、第二の四面体付加が起こって、セリンのヒドロキシ基が再生する。そして、プロトンが解放され、同時にタンパク質が新たにできたC末端と結合する[1]。